Elijah Oyoo-Okoth
Email: elijaoyoo2009@gmail.com
© 2019 Sift Desk Journals. All Rights Reserved
VOLUME: 3 ISSUE: 2
Page No: 222-231
Elijah Oyoo-Okoth
Email: elijaoyoo2009@gmail.com
Anthony Muthoni Mwangia, Charles, C. Ngugib, Jumbe James Jumbec, Elijah Oyoo-Okothd,*
aSchool of Pure and Applied Sciences, Kenyatta University
bSchool of Natural Resources and Environmental Studies, Karatina University, P.O. Box 1957-10101,
Karatina, Kenya
cDepartment of Zoological Sciences, Kenyatta University
dSchool of Environmental Studies, Department of Environmental Biology, University of Eldoret, P.O. Box 1125-30100, Eldoret–Kenya
A.M. Mwangi, C C. Ngugi, J.J.Jumbe, E.O. Okoth, Grading frequency affect the growth performance and intra−cohort cannibalism in African catfish (Clarias gariepinus, burchell, 1822) culture(2020)Journal of Aquaculture, Fisheries & Fish Science 3(2) pp:222-231
Growth of African catfish (Clarias garipinus) is characterized by large size heterogeneity resulting in poor growth performance and cannibalism. Strategies that limit size heterogeneity in C. gariepinus culture are therefore advocated. This study determined the effects of grading frequency on the growth performance and cannibalism of C. gariepinus in tank culture system. Three treatments designated as G0 (no grading), G2 (grading every 2 weeks) and G4 (grading every four weeks) in a Completely Randomized Design (CRD) was applied in triplicate. The tanks were stocked with 200 fingerlings each. Sampling was conducted weekly to measure length and weight of fish. Mortalities were recorded and dead fish observed under dissecting microscope to ascertain that it is due to cannibalism. Growth performance in terms of final mean weight, weight gain, specific growth rate and food conversion ratio were significantly (P < 0.05) affected by frequency of fish grading. The C. gariepinus graded every two weeks grew better than those graded every four weeks and were all above the no grading treatment. Significant changes in fish heterogeneity were observed after day 42. The size variation was significantly affected by grading frequency. The mean TL of fish graded every 2 weeks was the highest, followed by grading every four weeks and finally the no grading treatment was the lowest. Fish size heterogeneity (CV% and skewness) was consistently the highest in C. gariepinus in no grading treatment followed by fish where grading was done every four weeks while it was lowest in treatment where feeding was done every two weeks. Mortality owing to cannibalism was affected by grading frequency where highest cannibalism mortality occurred in C. gariepinus where there was no grading followed by grading every four weeks and intermediate at grading frequency every four weeks. Meanwhile other mortality by other causes such as such as wounds and suffocation in fish did not differ with grading frequency.
Keywords: Cannibalism, Coefficient of variation, Fish grading, Fish growth, Growth heterogeneity
Catfishes constitute a large group of freshwater fishes which are widely distributed throughout the world (Teugels, 1996). Clarias gariepinus is an important tropical catfish species for aquaculture, has an almost Pan−African distribution, ranging from the Nile to West Africa and from Algeria to Southern Africa (Brummett, 2008). Culture performance of C. gariepinus is well documented in several aquaculture production systems. The species is highly preferred due to its resistance to diseases, high fecundity and easy larvicidal production in captivity (Akinwole & Faturoti, 2007; Fauji et al., 2018; Huisman & Richter, 1987). The species is highly cultured in developing countries due to the additional qualities such as hardiness, ability to survive in different culture systems and diverse environments, and adaptation to supplemental feeds (Musa et al., 2012; Opiyo et al., 2017).
The C. garipinus exhibits allometric growth patterns where larger fish have higher growth potentials than small sized individuals (Ayo-Olalusi, 2014; Davies et al., 2013; Ekelemu, 2010). The size heterogeneity in young fish cohorts may affect overall on growth performance (Al-Hafedh & Ali, 2004; Martins et al., 2005a) or increase cannibalism in fish cohorts (Baras & Fortuné dAlmeida, 2001). The intensity of cannibalism would reach a maximum in the early weeks or months of the life history when the variability of individual growth is optimal (Baras & Jobling, 2002). Heterogeneous size distributions often lead to social dominance, which in turn results in aggressive behaviour and cannibalistic responses (Baras & Jobling, 2002). This in turn increases their susceptibility to diseases and weakens the fishes making them more liable to cannibalism or death (Baras & Fortuné dAlmeida, 2001; Hecht & Appelbaum, 1988; Hecht & Pienaar, 1993). Therefore, to enhance profitability from aquaculture production, strategies that are designed to improve growth performance and survival as well as reduce cannibalism should be explored.
Size grading practices is a common procedure used during intensive fish rearing to reduce size variations (Blonk et al., 2010; Heikes, 2003; Torrans & Ott, 2018), and has resulted in differences on growth, production, feed conversion, and food fish size distribution (Torrans & Ott, 2018). This practice assumes that larger individuals suppress the growth of smaller, subordinate fish. Moreover, size grading practices have been advocated method to control large differences in size of fish during the nursery period of many piscivorous fish species (Baras & Jobling, 2002). Sorting also optimizes feeding, since granulation and ration sizes can be tailored to the fish size (Batzina et al., 2018). The C. gariepinus is highly cannibalistic at the early larval stage (Baras, 2013) but knowledge of the changes in the species age and cannibalism is still scanty. Consequently, there is lack of information on the effect of age-based frequency of grading on growth performance, survival and cannibalism in C. gariepinus. Therefore, the aim of the current study was to determine the effects of size grading on the growth performance and cannibalism of C. gariepinus culture.
2.1 Experimental facility
The study was conducted in controlled laboratory conditions at Mwea Aquafish Farm Limited. The fish farm is located 110 km North East of Nairobi on the Nairobi-Embu highway and 1.5 km from Kimbimbi Town in Kirinyaga County, Kenya at a 0˚36.73’S, 37˚22.84’E and 1208 m above the sea level. Daily temperatures ranges from 16.9˚C−28˚C with cool seasons ranging between 12.9−23˚C and warm seasons between 18−28˚C. The culture condition were circular plastic tanks each with a surface area of 1.49 m2 and a height of 0.8 m supplied with 800 L of filtered dechlorinated tap water constantly regulated at a rate of approximately 50 L h–1. Tap water was stocked in an intermediate tank and aerated for at least 48 hours to remove chlorine before using it in the rearing facilities. To ensure that bacteria are destroyed in all aquaria, tap water used was treated with germicide UV lamp. The water was continuously aerated, and temperature ranges were 22.0−26°C. Salinity of the water in tanks (determined by salinometer, Model IC/SB–1 Salinity Cell) was 0.2 psu; NO2– < 0.05 mg L–1; NO3–1 <0.01 mg L–1; NH3 < 0.02 mg L–1; pH 6.7–73). The photoperiod regime was natural light of 12−h light and 12−h. Dissolved Oxygen (DO) was monitored every morning and found to range from 6−8 mg l−1.
2.2 Experimental fish
Four mature female broodstock (mean weight = 379.5 ± 24.5 g) and three mature males (mean weight = 360.9 ± 18.4 g) were obtained from the broodstock ponds within the facility and transferred to the hatchery at the Fish Farm. All African catfish parents conditioning, pituitary extraction, egg fertilization and incubation followed protocols as detailed by elsewhere (de Graaf & Janssen, 1996). Ambient water temperature in the incubator was maintained at 27 ± 0.1°C using a thermostat heater. Hatching began 24 hours after egg incubation. After complete hatch and yolk absorption, 100 larvae with a mean weight of 3.5 ± 0.1 mg) were siphoned out of the incubation tank and transferred into 27 glass aquaria each measuring 35 cm × 30 cm × 30 cm and containing 20 litres of water (5 larvae per litre) in a recirculation water system aerated with electric pump with air stone diffusers. Larvae were obtained through induced breeding and semi-natural spawning. The larvae were cultured for a period of 21 days to an initial mean weight of 26.3 ± 2.4 g.
2.3 Experimental design and setup
The study consisted of three treatments in a completely randomized design (CRD). The three treatments were designated as G0 (no grading), G2 (grading every 2 weeks) and G4 (grading every four weeks). The entire experiment was executed in triplicate. A total of 1800 fingerlings of the same cohort with a mean weight of 0.51 ± 0.12 g were transferred from the hatchery, where they were under acclimation and stocked in nine circular plastic tanks. The tanks were stocked with 200 fingerlings each. Fingerlings were fed with a diet containing 40% crude protein, 9% crude lipids, 3.5% crude fibre and 5.5% ash. The fish were manually hand fed at 8% body weight for the first month and then reduced to 6% body weight for the second month. Feed was administered four times per day i.e. at 0900 h, 1200 h, 0200 h and 0400 h, seven days a week. Tanks were cleaned twice a day to remove uneaten feed, faeces and dead fish. Formulated feed, water quality parameters and tank management were like the period of acclimation.
Grading was done by separating the jumpers/shooters from the rest of the fish population. Tanks were completely drained and observation made on presence of shooters upon which they were cropped out into separate holding tanks. Data on the number of shooters harvested from each tank was then recorded.
2.4 Fish Sampling, mortality and cannibalism
Sampling was conducted weekly. During sampling, dip nets were used to harvest 30 fish from each tank. Length and weight measurements fish was done from a randomly selected sample, n = 30. The wet weights were measured by use of a Sartorius Analytical Balance (readability 0.01 mg) and total length by use of a plastic ruler to the nearest 0.1 mm.
Mortalities were recorded every feeding session (0800 and 1600 hrs). Every dead fish was carefully observed under dissecting microscope to ascertain that it is not due to cannibalism. Final number of fish in each tank was recorded at the end of the culture period to determine the average survival.
Most of the visual observations of fingerling aggression and cannibalism were done between the periods of 0700 and 1900 hours during the 8−week period. Dead or eaten fish were not replaced during the experiment and cannibalism was calculated by recording the difference in fish numbers between the initial count and the final count of fish observed under dissecting microscope and cannibalism ascertained. Naturally occurring mortality was distinguished from mortality due to cannibalism and each parameter separately tallied for the tanks.
2.5 Water quality analyses and parameters
Water quality analyses was conducted to ensure that growth of fish is under the optimum water quality parameters and is not in any way influenced by the extreme’s long column sampler. Temperature was measured using a constant temperature meter while pH was measured using pH meter (Model: HI 98127; Manufacturer Hanna Instruments, USA). The DO was measured using DO meter (Model: YSI 550A; Manufacturer: Yellow Spring Instrument Company, Ohio, USA). Water samples were collected daily at 0800 h and 1800 h from the tanks using a 1.12 m and portions of the water samples used to determine: nitrate-nitrogen (NO3−N), total ammonia nitrogen (TAN), and soluble reactive phosphorus (SRP). All the analyses will be conducted following the standard analytical procedures (Bhatnagar & Devi, 2013). Water quality parameters were as follows: water temperature 26.0 ± 0.2°C (mean ± SD), pH 6.7−7.2, dissolved oxygen 5.4 ± 0.4 mg L−1, ammonia and nitrite nitrogen <0.6 mg L−1 and SRP ranged between 0.6 to 0.9 mg L−1.
2.6 Calculation of growth, mortality and cannibalism
In estimation of growth and survival for each experiment, about 20 fish in each tank were sampled fortnightly and weighed to calculate the individual mean weight and the following parameter calculated:
Weight gain = Initial Weight−Final weight
Percent weight gain = (Initial Weight−Final weight)/Initial weight×100
Specific growth rate (SGR, % day−1) = (eg – 1)100 where g = (ln(W2) − (ln(W1))(t2−t1)−1 and W2 and W1 are weights on day t2 and t1 respectively.
Survival = Final remaining fish/Initial number stocked×100.
Cannibalism = (Number of fingerling missing or consumed/Initial number of fish) × 100.
FCR Feed intake/weight gain
2.7 Data analysis
Before statistical analysis, normality of the data was determined using Shapiro−Wilk test (Hanusz et al., 2016), while homogeneity of variance was ascertained using Levene’s test (Shear et al., 2018). Statistical analyses were done using SPSS version 23.0. The effect of grading frequency (factors) on growth performance and cannibalism were performed by analysis of variance (One-way ANOVA). Significantly different were compared using Duncans Multiple Range Test (DMRT). Values throughout the text are expressed as mean ± standard Error. In all the analysis significant was accepted at P < 0.05.
Growth performance and feed utilization of C. gariepinus in tanks under different grading frequencies are presented in Table 1. Clarias gariepinus graded every two weeks grew better than those graded every four weeks and were all above the no grading treatment. After 60 days of experiment, the final mean weight and weight gain in C. gariepinus among treatments were significantly (P < 0.05) the highest in treatment involving grading every two weeks while the lowest mean weight and weight gain being recorded in fish that were not graded (control). Specific growth rate (SGR) was significantly (P < 0.05) highest in fish that were graded every two weeks followed by fish graded every four weeks while no grading treatment produced the lowest mean weight and weight gain. The lowest FCR also occurred in treatment that were graded every two-week followed by those graded every four week and lowest in the no grading treatment (P < 0.05).
Table1: Growth performance (means ± SD) of C. gariepinus in different grading treatments
|
Growth performance parameters |
Frequency of grading |
||
|
G0 (No grading) |
G2 (Every 2 weeks) |
G4 (Every 4 weeks) |
|
|
Initial mean fish weight (g) |
0.69 ± 0.31 |
0.92 ± 0.39 |
0.89 ± 0.21 |
|
Final mean weight (g) |
36.32 ± 10.3a |
51.72 ± 11.15c |
42.48 ± 8.2b |
|
Weight gain (g) |
35.63 ± 10.02a |
50.8 ± 12.42c |
41.59 ± 10.27b |
|
Specific growth rate (SGR; % day−1) |
8.81 ± 0.27b |
8.95 ± 0.08c |
8.59 ± 0.06b |
|
FCR |
1.81 ± 0.23c |
1.21 ± 0.13a |
1.42 ± 0.24b |
Changes in weight of the fish under different grading treatments at the start (day 28), day 42, day 56 and day 60 are shown in Fig. 1. There were no significant differences in grading treatment on day 28 (P > 0.05). However, the differences in the weight due to grading occurred from day 42 to 60 (P < 0.05) where grading every two weeks produced the best weight followed by grading every four weeks and low weight in no grading treatments.
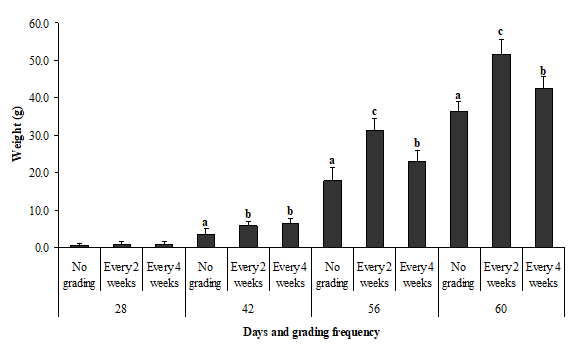
Figure 1: Changes in weight of the fish under different grading treatments at the start (day 28), day 28, day 56 and day 60. Different low-case letters as superscript within the same day represent significant differences in growth between grading frequencies.
Results showing effects of frequency of grading on the total length (TL, cm) at day 60 (when there were large size variations) is shown in Fig. 2. The size variation was significantly affected by grading frequency (P < 0.05). The mean TL of fish graded every 2 weeks was the highest (19.1 ± 2.9 cm), followed by grading every four weeks (17.4 ± 2.3 cm) and finally the no grading treatment was the lowest (15.8 ± 1.9 cm).
Fish size heterogeneity (CV, TL and skewness) of different grading treatments in Clarias gariepinus is shown in Fig. 3. Fish size heterogeneity (CV%) was consistently the highest in C. gariepinus in no grading treatment (21.3 ± 3.2%) followed by fish where grading was done every four weeks (13.4± 2.2%) while it was lowest in treatment where feeding was done every two week (7.4± 1.3%). Fish size skewness followed the patterns of CV where it was consistently skewed in C. gariepinus in no grading treatment followed by f grading every four weeks while least skew was obtained in treatment where feeding was done every two weeks.
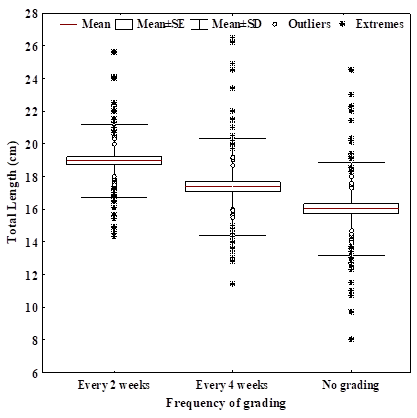
Figure 2: Fish Total Length (TL) at different grading treatments in Clarias gariepinus
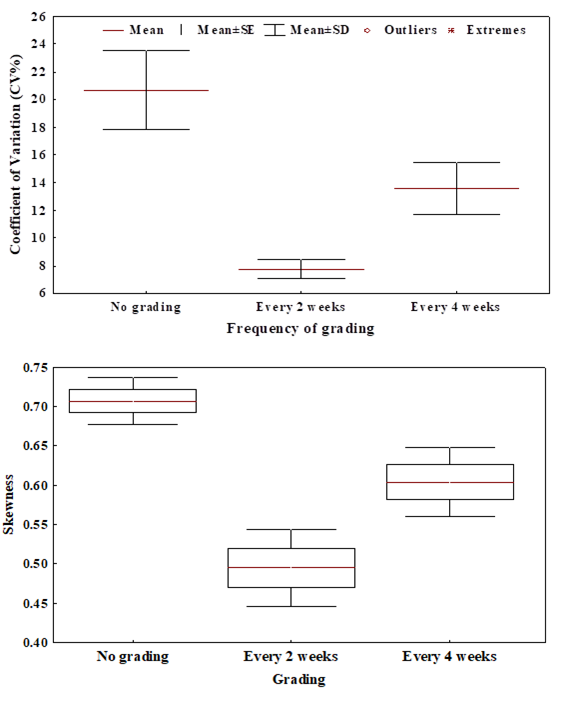
Figure 3: Fish size heterogeneity (CV, TL and skewness) of different grading treatments in Clarias gariepinus
Table 2: Mortality due to cannibalism and other causes in fish under different frequency of grading
|
Grading frequency |
Cannibalism mortality (%) |
Mortality by other causes (%) |
Total mortality (%) |
|
G0 (No grading) |
42.4 ± 10.2c |
11.5 ± 12.3 |
56.8 ± 15.3c |
|
G2 (Every 2 weeks) |
7.5 ± 2.0a |
10.8 ± 3.1 |
18.4 ± 4.4a |
|
G4 (Every 4 weeks) |
18.9 ± 4.9b |
13.7 ± 3.2 |
35.6 ± 14.5 b |
|
One-Way ANOVA |
|
|
|
|
F |
23.4522 |
3.3212 |
21.3342 |
|
P value |
0.0000 |
0.09222 |
0.0000 |
Mortality owing to cannibalism was significantly affected by grading frequency (P < 0.001, Table 2). Highest cannibalism mortality was highest in C. gariepinus where there was no grading followed by grading every four weeks and intermediate at grading frequency every four weeks. Meanwhile other mortality by other causes such as such as wounds and suffocation in fish did not differ with grading frequency at al. As a result, there were significant differences in total mortality of fish based in differential grading frequency (P < 0.001) where grading every two weeks resulted in the lowest total mortality.
In this study, the growth rates were significantly affected by frequency of grading. Clarias gariepinus graded every two weeks grew better than those graded every four weeks and were all above the no grading treatment with highest final mean weight and weight gain, SGR and lowest FCR occurring the grading every two weeks treatment. Growth rate was improved by 15% and 12% in the grading every two weeks and grading every four weeks over the control group. This is in agreement with previous studies that have established that size grading affect the growth performance of several species of fish (Barki et al., 2000; Kamstra, 1993; Lambert & Dutil, 2001; Mgaya & Mercer, 1995; Mun et al., 2019; Nightingale et al., 2018). In restricted spaces such as ponds, aquaria and concrete tanks, the African catfish tends to exhibit changes in behavioural patterns when there are size variations in the cohorts leading to reduced feed intake, aggression, death and cannibalism (Martins et al., 2005b). Accordingly, several proposals to conduct size grading of juvenile C. gariepinus at regular basis have been suggested but the regularity of the grading is rarely studied. In this study grading every two weeks led to the best growth in such groups compared with rearing the fish in groups with larger initial variation in weight. A lower growth in juvenile halibut has been reported when reared with similar sized fish compared with groups where medium sized fish were reared with either larger conspecifics (defined as dominates) or smaller conspecifics (defined as subordinates) (Stefánsson et al., 2000). Some mechanisms have been proposed to explain how grading of fish influence the growth, e.g., physiological stress (Griffiths & Armstrong, 2002; Kestemont et al., 2003), disproportional food acquisition (Azaza et al., 2013; Barbosa et al., 2006), and activity differences (Sloman & Armstrong, 2002). Relative size of fish will be an important determinant of fighting ability in aggressive behavior as well as attacking opponents and obtaining preferential access to food.
Most of the size heterogeneity occurred after 42 days of experiment. Cannibalism is size restricted in the early life stages due to gape size limitation (Nilsson & Brönmark, 2000). Cannibalistic fish are morphologically capable of ingesting a maximum conspecific prey up to 78% of their body length (Ribeiro & Qin, 2013), but the predation efficiency is negatively correlated to prey size as prey smaller than 50% of the cannibal size can result in high energetic return to the cannibal (Ribeiro & Qin, 2015). Therefore, once the 50% threshold of prey–cannibal size difference was attained, potential cannibals emerged and their relative abundance dictated the severity of the upcoming cannibalism mortality.
The current findings indicate that individual C. gariepinus grow better when they are reared in uniform sized grading cohots. The data show better growth of fish graded every two weeks and hence low size heterogeneity compared with ungraded. One possible explanation might be a higher level of intraspecific competition and agonistic interaction in similar sized groups which have been graded every two weeks. Another possible explanation of the positive growth effects could be due to the removal of possible dominates and subordinates. The term ‘‘resource-dependent competition’’ describe the process where resource competition, e.g. for food, controls the relative growth rates and there are cases in which size hierarchies develop in the presence of excess food. It was suggested that size hierarchy (with larger fish dominating smaller ones) forms where the dominant fish consumes a greater proportion of the group meal (McCarthy et al. 1992; Jobling 1995). Less resource−dependent competition after grading would explain the positive effect seen in the SL group in this study. In the case of the LS group, removal of subordinates could change the social structure within the group and lead to less intraspecific competition, thus allowing the larger fish to fulfil their growth potential. The findings of Imsland et al. (1998) support this explanation as indications of size-dependency in formations of size hierarchies, because different forms of size hierarchy were seen within three grading groups (large, medium, small). For the fish not graded there may be more disproportionate interactions of fish within the same social hierarchy than in fish graded every two weeks where the size heterogeneity was low. Therefore, grading every two weeks may have reduced intra-specific interactions with possible positive effects on growth rate. Hence, both scenarios (dominate suppression and subordinate competition) are supported by the current findings. This could indicate a stronger social hierarchy and a higher level of intraspecific competition in the former groups, with a possible negative effect on growth.
The formation of size hierarchies in fish is generally found to have negative effects on overall biomass gain, leading to inconsistency of growth among groups and heterogeneity in growth among individuals within a group (Jobling and Koskela 1996; Stefa´nsson et al. 2000). In this study, the size variation, and heterogeneity were significantly affected by grading frequency where mean TL, CV and skewness of fish graded every 2 weeks was the highest followed by grading every four weeks were all above the control. It is generally accepted that intraspecific competition and agonistic interaction leading to size heterogeneity and cannibalism are found to be greater when fish of different size are reared together in fishes like Arctic charr Salvelinus alpinus L. (Baardvik and Jobling, 1990), turbot Scophthalmus maximus (Sunde et al., 1998) and Atlantic halibut Hippoglossus(Stefánsson et al., 2000). An interesting observation in early juvenile African catfish is that as food availability and acceptability starts to decline, territoriality and aggressive behaviour appears to increase. In the first two weeks of the study, there was a failure of the catfish fingerlings to accept the formulated feed. This phenomenon might have resulted in a semi-starvation situation which probably accounted for the recorded cannibalism. Two forms of cannibalism were observed in study period; Group cannibalism, where individuals of approximately the same size attacked and consumed a weaker, injured or dead fingerling and complete ingestion, where a larger individual wholly swallowed smaller fingerlings, however, during the recording all these were included as cannibalism mortality. Imsland et al. (1998) used individual-based models to investigate the causes of size variation in a culture population of juvenile turbot. Two variants of size hierarchies were included in their models. One variant (A) was a dominance hierarchy in which every fish is subordinate to a larger individual; the other (B) was size-related dominance into dominant (the largest fish) and subordinates (the smallest fish). The study of Imsland et al. (2013) showed a difference in the sensitivity of the two types of size hierarchy tested in different grading groups at different times, indicating formation of different types of size hierarchy in different size groups in juvenile turbot. They concluded that social interactions related to size-dependent hierarchies contribute to size variation in juvenile turbot. In the current study the treatment being graded every two weeks displayed the least size variation.
Mortality owing to cannibalism was significantly affected by grading frequency. The highest mortality due to cannibalism occurred in no grading treatment followed by grading every four weeks and intermediate at grading frequency every four weeks. The first incident of cannibalism was observed during the second week in tank 2. Meanwhile other mortality by other causes such as such as wounds and suffocation in fish did not differ with grading frequency at al. As a result, there were significant differences in total mortality of fish based in differential grading frequency where grading every two weeks resulted in the lowest total mortality. Over the 60 days period, cannibalism accounted for 42.4% of the total juvenile catfish population for the study where there was no grading done. This represented a significant proportion of the initial catfish population and might have been largely influenced by the limited spaces of the aquaria. The size disparity between the relatively larger fingerling that exhibited the abnormal growth spurt and the other fingerlings, however accounted for the higher rate of cannibalism in tank 2 and in the whole research in general. Abdelhamid et al. (2010) in their study on improving the survival rate of the African catfish, prescribed the grading technique of separating of the biggest fry from the general population as a means of reducing the cannibalism phenomena.
The growth performance in terms of final mean weight, weight gain, SGR and FCR in C. gariepinus was affected by frequency of fish grading. The C. gariepinus graded every two weeks grew better than those graded every four weeks and were all above the no grading treatment. Changes in fish heterogeneity was observed after day 42 and continued until day 60. The size variation was significantly affected by grading frequency. The mean TL of fish graded every 2 weeks was the highest, followed by grading every four weeks and finally the no grading treatment was the lowest. Fish size heterogeneity (CV% and skewness) was consistently the highest in C. gariepinus in no grading treatment followed by fish where grading was done every four weeks while it was lowest in treatment where feeding was done every two weeks. It was also established that mortality owing to cannibalism was affected by grading frequency where highest cannibalism mortality occurred in C. gariepinus where there was no grading followed by grading every four weeks and intermediate at grading frequency every four weeks. Meanwhile other mortality by other causes such as such as wounds and suffocation in fish did not differ with grading frequency.
Akinwole, A. & Faturoti, E. (2007). Biological performance of African Catfish (Clarias gariepinus) cultured in recirculating system in Ibadan. Aquacultural engineering, 36(1), 18-23.
View ArticleAl-Hafedh, Y. & Ali, S. (2004). Effects of feeding on survival, cannibalism, growth and feed conversion of African catfish, Clarias gariepinus (Burchell) in concrete tanks. Journal of Applied Ichthyology, 20(3), 225-227.
View ArticleAyo-Olalusi, C. (2014). Length-weight Relationship, Condition Factor and Sex Ratio of African Mud Catfish (Clarias gariepinus) Reared in Flow-through System Tanks. Journal of Fisheries and Aquatic Science, 9(5), 430-434.
View ArticleAzaza, M., Assad, A., Maghrbi, W. & El-Cafsi, M. (2013). The effects of rearing density on growth, size heterogeneity and inter-individual variation of feed intake in monosex male Nile tilapia Oreochromis niloticus L. animal, 7(11), 1865-1874. PMid:23915501
View Article PubMed/NCBIBaras, E. (2013). Cannibalism in fish larvae: what have we learned. Larval fish aquaculture, 167-199.
Baras, É. & Fortuné dAlmeida, A. (2001). Size heterogeneity prevails over kinship in shaping cannibalism among larvae of sharptooth catfish Clarias gariepinus. Aquatic Living Resources, 14(4), 251-256. 01118-4
View ArticleBaras, E. & Jobling, M. (2002). Dynamics of intracohort cannibalism in cultured fish. Aquaculture research, 33(7), 461-479.
View ArticleBarbosa, J., Brugiolo, S., Carolsfeld, J. & Leitao, S. (2006). Heterogeneous growth fingerlings of the Nile tilapia Oreochromis niloticus: effects of density and initial size variability. Brazilian Journal of Biology, 66(2A), 537-541. PMid:16862309
View Article PubMed/NCBIBarki, A., Harpaz, S., Hulata, G. & Karplus, I. (2000). Effects of larger fish and size grading on growth and size variation in fingerling silver perch. Aquaculture International, 8(5), 391-401.
View ArticleBatzina, A., Drossos, I. P. & Karakatsouli, N. (2018). Effects of grading on individual growth and feeding behaviour of European seabass Dicentrarchus labrax. Aquaculture research, 49(12), 3759-3768.
View ArticleBhatnagar, A. & Devi, P. (2013). Water quality guidelines for the management of pond fish culture. International journal of environmental sciences, 3(6), 1980-2009.
Blonk, R. J., Komen, H., Kamstra, A. & van Arendonk, J. A. (2010). Effects of grading on heritability estimates under commercial conditions: A case study with common sole, Solea solea. Aquaculture, 300(1-4), 43-49.
View ArticleBrummett, R. E. (2008). Clarias catfish: biology, ecology, distribution and biodiversity, Proceedings of a workshop on the development of a genetic improvement program for African catfish Clarias gariepinus. WorldFish Center conference proceedings, pp. 64-72.
Davies, O. A., Tawari, C. C. & Kwen, K.-I. (2013). Length-Weight Relationship, Condition factor and Sex ratio of Clarias gariepinus juveniles reared in concrete tanks. International Journal of Scientific Research in Environmental Sciences, 1(11), 324.
View Articlede Graaf, G. & Janssen, J. (1996). Handbook on the artificial reproduction and pond rearing of the African catfish Clarias gariepinus in sub-saharan Africa. FAO fisheries technical paper, 362, 13-50.
Ekelemu, J. K. (2010). Differential growth patterns of Clarias gariepinus, Heterobranchus bidorsalis and Hybrid Heteroclarias fed commercially prepared diets. Agric. Biol. JN Am, 1(4), 658-661.
Fauji, H., Budiardi, T. & Ekasari, J. (2018). Growth performance and robustness of African Catfish Clarias gariepinus (Burchell) in biofloc‐based nursery production with different stocking densities. Aquaculture Research, 49(3), 1339-1346.
View ArticleGriffiths, S. W. & Armstrong, J. (2002). Rearing conditions influence refuge use among over‐wintering Atlantic salmon juveniles. Journal of Fish Biology, 60(2), 363-369.
View ArticleHanusz, Z., Tarasinska, J. & Zielinski, W. (2016). Shapiro-Wilk test with known mean. REVSTAT-Statistical Journal, 14(1), 89-100.
Hecht, T. & Appelbaum, S. (1988). Observations on intraspecific aggression and coeval sibling cannibalism by larval and juvenile Claias gariepinus (Clariidae: Pisces) under controlled conditions. Journal of Zoology, 214(1), 21-44.
View ArticleHecht, T. & Pienaar, A. G. (1993). A review of cannibalism and its implications in fish larviculture. Journal of the World Aquaculture Society, 24(2), 246-261.
View ArticleHeikes, D. (2003). Development of In-Pond Grading Technology for Catfish and Hybrid Striped Bass. Educational Program s 57.
Huisman, E. & Richter, C. (1987). Reproduction, growth, health control and aquacultural potential of the African catfish, Clarias gariepinus (Burchell 1822). Aquaculture, 63(1-4), 1-14. 90057-3
View ArticleKamstra, A. (1993). The effect of size grading on individual growth in eel, Anguilla anguilla, measured by individual marking. Aquaculture, 112(1), 67-77. 90159-V
View ArticleKestemont, P. et al. (2003). Size heterogeneity, cannibalism and competition in cultured predatory fish larvae: biotic and abiotic influences. Aquaculture, 227(1-4), 333-356. 00513-1
View ArticleLambert, Y. & Dutil, J.-D. (2001). Food intake and growth of adult Atlantic cod (Gadus morhua L.) reared under different conditions of stocking density, feeding frequency and size-grading. Aquaculture, 192(2-4), 233-247. 00448-8
View ArticleMartins, C. I., Aanyu, M., Schrama, J. W. & Verreth, J. A. (2005a). Size distribution in African catfish (Clarias gariepinus) affects feeding behaviour but not growth. Aquaculture, 250(1-2), 300-307.
View ArticleMartins, C. I., Schrama, J. W. & Verreth, J. A. (2005b). Inherent variation in growth efficiency of African catfish Clarias gariepinus (Burchell, 1822) juveniles. Aquaculture Research, 36(9), 868-875.
View ArticleMgaya, Y. D. & Mercer, J. P. (1995). The effects of size grading and stocking density on growth performance of juvenile abalone, Haliotis tuberculata Linnaeus. Aquaculture, 136(3-4), 297-312. 00066-6
View ArticleMun, S. H. et al. (2019). Expression Patterns of Growth Related Genes in Juvenile Red Spotted Grouper (Epinephelus akaara) with Different Growth Performance after Size Grading. Development & reproduction, 23(1), 35. PMid:31049470
View Article PubMed/NCBIMusa, S. M., Aura, C. M., Ngugi, C. C. & Kundu, R. (2012). The effect of three different feed types on growth performance and survival of African catfish fry (Clarias gariepinus) reared in a hatchery. ISRN Zoology, 2012.
View ArticleNightingale, J., Stebbing, P., Taylor, N., McCabe, G. & Jones, G. (2018). Assessing the effect of size‐grading for rearing young‐of‐the‐year white‐clawed crayfish Austropotamobius pallipes. Aquaculture Research, 49(9), 3116-3122.
View ArticleNilsson, P. A. & Brönmark, C. (2000). Prey vulnerability to a gape‐size limited predator: behavioural and morphological impacts on northern pike piscivory. Oikos, 88(3), 539-546.
View ArticleOpiyo, M. A., Orina, P. & Charo-Karisa, H. (2017). Fecundity, growth parameters and survival rate of three African catfish (Clarias gariepinus) strains under hatchery condition.
View ArticleRibeiro, F. & Qin, J. (2015). Prey size selection and cannibalistic behaviour of juvenile barramundi Lates calcarifer. Journal of fish biology, 86(5), 1549-1566. PMid:25801794
View Article PubMed/NCBIRibeiro, F. F. & Qin, J. G. (2013). Modelling size-dependent cannibalism in barramundi Lates calcarifer: cannibalistic polyphenism and its implication to aquaculture. PloS one, 8(12). PMid:24349295
View Article PubMed/NCBIShear, B. R., Nordstokke, D. W. & Zumbo, B. D. (2018). A Note on Using the Nonparametric Levene Test When Population Means Are Unequal. Practical Assessment, Research, and Evaluation, 23(1), 13.
Sloman, K. A. & Armstrong, J. (2002). Physiological effects of dominance hierarchies: laboratory artefacts or natural phenomena? Journal of Fish Biology, 61(1), 1-23.
View ArticleStefánsson, M. et al. (2000). The effect of different initial size distributions on the growth of Atlantic halibut. Journal of fish biology, 56(4), 826-836.
View ArticleTeugels, G. G. (1996). Taxonomy, phylogeny and biogeography of catfishes (Ostariophysi, Siluroidei): an overview. Aquatic Living Resources, 9(S1), 9-34.
View ArticleTorrans, L. & Ott, B. (2018). Effect of grading fingerling hybrid catfish (♀ Channel Catfish×♂ Blue Catfish) on growth, production, feed conversion, and food fish size distribution. North American Journal of Aquaculture, 80(2), 187-192.
View Article