Mingqiang Qiao
Tel: +86-022-2350-3692; Fax: +86-022-2350-3340
E-mail: qiaomq@nankai.edu.cn
© 2019 Sift Desk Journals. All Rights Reserved
VOLUME: 4 ISSUE: 6
Page No: 815-821
Mingqiang Qiao
Tel: +86-022-2350-3692; Fax: +86-022-2350-3340
E-mail: qiaomq@nankai.edu.cn
Yuzhen Wu1, Quanli Liu1, Yiming Chen1, Haijin Xu1, Ossi Pastinen2, Ossi Turunen3, Mingqiang Qiao1*
1 The Key Laboratory of Molecular Microbiology and Technology, Ministry of Education, College of Life Sciences, Nankai University, Tianjin, 300071, China
2 Department of Bioproducts and Biosystems, School of Chemical Engineering, Aalto University, P.O. Box 16100, FI-00076 Aalto, Finland
3 Faculty of Science and Forestry, School of Forest Sciences, University of Eastern Finland, P.O. Box 111, FI-80101 Joensuu, Finland
Liang Guo(zsdxgl@163.com)
Xiuping He(hexp@im.ac.cn)
Gang Liu(liug@im.ac.cn)
Zihe Liu(zihe@mail.buct.edu.cn)
Mingqiang Qiao, Yuzhen Wu, Quanli Liu, Yiming Chen, Haijin Xu, Ossi Pastinen, Ossi Turunen, Large genome deletions reveal gene effects on ethanol tolerance in Saccharomyces cerevisiae (2019) Journal of Food Science & Technology 4(6)
The yeast Saccharomyces cerevisiae, which is a model system for biological research, has been widely used as an industrial host. Using the mazF-mediated deletion system, we made large deletions, including both terminal and internal regions of the chromosome. We obtained 12 deletion strains with different sizes of the deleted genome region(s), varying from 20 kb to 80 kb. Each deletion removed a cluster of non-essential genes, as deduced by an online prediction program. Phenotype analysis of the mutants with large deletions showed that genome regions YOR124C/YOR134W and YOL020W/YOL011W appeared to be related to ethanol tolerance and high salinity sensitivity, respectively. The loss of CAT5, ADE2, or RGA1 reduced the ethanol tolerance of yeast. Dispensable genes or interactions among non-essential genes were involved in the functional effects of genome deletions.
Key words: ethanol tolerance; genome deletions; non-essential genes; oxygen response; Saccharomyces cerevisiae
The present-day genome of the yeast Saccharomyces cerevisiae was formed by whole genome duplication, after which the genome evolved because of the loss of a large part of the duplicated genes and asymmetric divergence of at least part of the remaining duplicated genes [1-3]. Yeast genome has functioned as a significant model to study the eukaryotic genetic system [3], and it contains a proportion of redundant chromosomal sequences. Recent studies showed great interest in the streamlining of genome sequence to obtain cells with optimized characteristics. In Escherichia coli, genome-reduced strains showed better growth fitness [4] and a 2.4-fold increase in L-threonine production as a production host [5]. In Bacillus subtilis, the strains with specific gene modifications related to guanosine and thymidine metabolism, as well as large genome reduction, improved the properties needed to produce nucleoside products [6]. Murakami et al. constructed a yeast mutant that had lost about 5% of the genome size though the chromosome-splitting techniques, and the mutant showed increased ethanol and glycerol production without weakening of its resistance under various stress conditions [7].
The budding yeast has been widely used for fermentation in industrial production. During fermentation, yeast cells may be exposed to extreme conditions, such as heat shock, hyperosmotic stress, pH changes, and increased ethanol concentration, which requires enhancement of tolerance to these kinds of stress factors. Ethanol tolerance was significantly enhanced by the deletion of valine accumulation gene LEU4 and LEU9 or INM1and INM2, which resulted in the reduction of inositol levels, as studied via a semi-rational approach based on yeast cell metabolomics [8]. By screening a transposon-mediated mutant library, Park et al. found that salt stress tolerance can be conferred with the open reading frame disruption of genes MDJ1 and VPS74 [9]. Except for these metabolic engineering strategies, the large genome region modifications could also generate strains that can produce the desirable chemicals at high concentrations.
Murakami et al. deleted portions of the yeast genome via chromosome-splitting method, which is efficient for large deletions, but the target DNA is limited to terminal chromosome regions [7]. The mazF-mediated deletion method [10], an optimized version with a strong counter selective marker mazF of Latour system [11] was efficient and suitable for both terminal and internal chromosome regions. Furthermore, this method relies solely upon homologous recombination and shows extremely good stability when it comes to different targets. In this study, we obtained 12 strains with deleted genome regions and evaluated their growth and stress response under different conditions. Region YOR124C/YOR134W and YOL020W/YOL011W were related to ethanol tolerance and high salinity tolerance, respectively. With region YOR124C/YOR134W as example, we further analyzed the specific genes included in this genome region and ADE2, CAT5, and RGA1 genes were needed for higher ethanol tolerance.
Strains and culture conditions
The large genome reduction strains and single gene knock-out strains used in this study are shown in Tables 1 and 2, respectively. All genome deletion strains are derived from the wild-type strain BY4741 (MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0). Yeast strains were cultured at 30 °C in YPD rich medium or in synthetic complete (SC) medium lacking relevant auxotrophic components, unless otherwise stated. SG medium was used to induce the expression of mazF by replacing glucose with galactose in SC medium. E. coli DH5α used for plasmid recovery was grown in LB medium at 37 °C.
Table 1. Genome reduced strains in this study
|
Name |
Deleted region |
Deletion size (kbp) |
No. of genes |
Source |
|
NK11 |
YKL072W/YKL061W |
26.5 |
12 |
[1] |
|
NK12 |
YKL224C/YKL215C |
28.8 |
10 |
[1] |
|
NK32 |
YIL014W/YIL005W |
21.7 |
11 |
This study |
|
NK33 |
YOR124C/YOR134W |
24.9 |
11 |
This study |
|
NK34 |
YOL020W/YOL011W |
21.2 |
11 |
This study |
|
NK13 |
YKL072W/YKL061W YKL224C/YKL215C |
55.3 |
22 |
This study |
|
NK42 |
YKL072W/YKL061W YOL020W/YOL011W |
47.7 |
23 |
This study |
|
NK43 |
YKL224C/YKL215C YOL020W/YOL011W |
50 |
21 |
This study |
|
NK38 |
YKL224C/YKL215C YIL014W/YIL005W |
50.5 |
21 |
This study |
|
NK39 |
YKL072W/YKL061W YKL224C/YKL215C YIL014W/YIL005W |
77 |
33 |
This study |
|
NK44 |
YKL224C/YKL215C YOL020W/YOL011W YIL014W/YIL005W |
71.7 |
32 |
This study |
|
NK45 |
YKL072W/YKL061W YKL224C/YKL215C YOR124C/YOR134W |
80.2 |
33 |
This study |
Table 2. Strains with single gene knock-out used in this study
|
No. |
Deleted gene |
Gene name |
Source |
|
H1 |
YOR124C |
UBP2 |
[2] |
|
H2 |
YOR125C |
CAT5 |
[2] |
|
H3 |
YOR126C |
IAH1 |
[2] |
|
H4 |
YOR127W |
RGA1 |
This study |
|
H5 |
YOR128C |
ADE2 |
This study |
|
H6 |
YOR129C |
AFI1 |
[2] |
|
H7 |
YOR130C |
ORT1 |
[2] |
|
H8 |
YOR131C |
|
[2] |
|
H9 |
YOR132W |
VPS17 |
[2] |
|
H10 |
YOR133W |
EFT1 |
[2] |
|
H11 |
YOR134W |
BAG7 |
[2] |
|
B1 |
YOL011W |
PLB3 |
[2] |
|
B2 |
YOL012C |
HTZ1 |
[2] |
|
B3 |
YOL013C |
HRD1 |
[2] |
|
B4 |
YOL013W-A |
|
[2] |
|
B5 |
YOL013W-B |
|
[2] |
|
B6 |
YOL014W |
|
[2] |
|
B7 |
YOL015W |
IRC10 |
[2] |
|
B8 |
YOL016C |
CMK2 |
[2] |
|
B9 |
YOL017W |
ESC8 |
[2] |
|
B10 |
YOL018C |
TLG2 |
[2] |
|
B11 |
YOL019W |
|
[2] |
|
B12 |
YOL019W-A |
|
[2] |
|
B13 |
YOL020W |
TAT2 |
[2] |
Construction of genome region(s) deletion strains and single gene knock-out strains
Candidate regions were selected by using the online program (http://gbpr.riise.hiroshima-u.ac.jp/index_en.php) for prediction of deletable regions in the yeast genome based on the datasets in the SGD and CYGD [7]. For the large chromosomal deletions, we used the mazF-mediated deletion system comprising two steps of homologous recombination. The details of the deletion system have been reported earlier [10]. The first step was the co-transformation of the flanking segments HA1, HA2, HA3, selection marker URA3 or LEU2, and counter selection marker mazF. We obtained a latent strain with the deletion cassettes integrated into the upstream of the target sequence through homologous recombination with the help of 50 bp sequence overlap. In the second step, the latent strain was spread on the SG medium to excise the target sequence and markers, and the transformants were verified by colony PCR.
Gene knock-out strains H1 to H11 and B1 to B13 were given by Professor Yingjin Yuan (Tianjin University, China), except for H4 (rga1Δ) and H5 (ade2Δ), which were constructed through homologous recombination by replacing the ORFs with selective marker in this study [12]. The URA3 and LEU2 markers were amplified by PCR by using pUG72 and pUG73 as templates, respectively. For gene complementation, RGA1 was inserted into the original genome locus with a selective marker. Two homologies corresponding to the 5’ and 3’ends of integration locus, RGA1 gene, and selective marker were obtained by PCR using genome DNA and pUG73 as templates. These fragments were assembled into deletion cassettes through homologous recombination among the 50 bp sequence overlaps by co-transformation. The cat5 and ade2 mutant cells were transformed with plasmids containing the wild-type CAT5, ADE2, or an empty vector.
Analysis of growth profile and stress tolerance
Inoculation cultures were obtained from cultures that had been inoculated from loop-fulls of cells from agar plates and incubated overnight (approximately 16 h) at 30 °C on a rotator in 5 ml YPD medium in 15 ml test tubes. Pre-cultures were harvested by centrifugation (500×g, 5 min, room temperature), washed twice in 5 ml sterile MilliQ water, and adjusted to a cell density with OD600 of 1.0 (approximately 1×107 cells ml-1) in fresh YPD medium.
For aerobic growth measurements, pre-cultures were inoculated to an OD of 0.1 in 50 ml flasks with 20 ml medium in each culture. The flasks were kept at 30 °C, and the cultures were subjected to orbital shaking at high intensity (220 rpm). Anaerobic growth curve was measured using 15 ml plastic tubes with caps filled with medium. Optical density measurements were manually performed at 8 h time intervals at 600 nm in a spectrophotometer.
For the assay of yeast growth under various stress factors, serial tenfold dilutions of the described pre-cultures were spotted onto a solid YPD media, and plates were incubated at 30 °C for 3 days. Concentrations of used inhibitors were as follows: ethanol, 8%; NaCl, 1M; and sorbitol, 1.5 M. For heat-shock response, the cell suspension was incubated at 50 °C for 30 min and cooled to 30°C, followed by spotting on YPD plate. Alkaline (pH 8.5) media were prepared by adding 0.1 M Tris·HCl buffer, pH 8.5, to YPD medium.
Construction of plasmids for gene complementation
The pLC42 plasmid was constructed by changing the selective marker to LEU2 in pSP-G1 cutting with Nde I (Takara). Both the pSP-G1 and pLC42 plasmid were used as control while gene complementation. pNC3 and pNC4 were constructed with pLC42 backbone cut with Sac II and Sma I and the backbone was gel-purified (Takara). The CAT5 and ADE2 sequence was PCR amplified with two primers containing the enzyme recognition site. The PCR product was then digested and purified. Purified fragments were combined using T4 DNA ligation (Thermo Fisher Scientific), and the resulting plasmids pNC3 and pNC4 were confirmed via restriction analysis. Both pNC4 and pSP-G1 were digested by Spe I and Not I, and the purified fragments were ligated by T4 DNA ligation, thereby generating plasmid pNC6. Plasmids used in this study are shown in Table 3.
Table 3. The plasmids used in this study
|
Name |
Characteristics |
Source or reference |
|
|
pUG72 |
PCR template for URA3 amplification |
[3] |
|
|
pUG73 |
PCR template for LEU2 amplification |
[3] |
|
|
pLC2 |
PCR template for mazF amplification |
[1] |
|
|
pSP-G1 |
2 μm ori, URA3, PTEF1- TADH1, PPGK1-TCYC1, Ampr |
[4] |
|
|
pLC42 |
2 μm ori, LEU2, PTEF1- TADH1, PPGK1-TCYC1, Ampr |
[5] |
|
|
pNC3 |
pLC42:: PTEF1- TADH1, PPGK1-CAT5-TCYC1 |
This study |
|
|
pNC4 |
pLC42:: PTEF1- TADH1, PPGK1-ADE2-TCYC1 |
This study |
|
|
pNC5 |
pSP-G1:: PTEF1-ADE2- TADH1, PPGK1-TCYC1 |
This study |
|
Molecular biology techniques
PCR amplification was carried out with I-5™ 2× High-Fidelity Master Mix (MCLAB) according to the manufacturer’s instructions. Colony PCR was performed with the SapphireAmp® Fast PCR Master Mix (Takara). Primers used in this study can be found in Table S1 (Supplementary data). The Plasmids and PCR fragments were digested with restriction enzymes and from Takara. Plasmids and yeast genomic DNA were isolated with TIANprep Mini Plasmid Kit (TIANGEN, Beijing, China) and EasyPure Genomic DNA Kit (TransGen Biotech, Beijing, China), respectively. Yeast competent cell preparation and yeast transformation were performed using the lithium acetate protocol [13].
Construction of strains with genome region(s) deleted
Five chromosome regions with more than 10 continuous non-essential genes were randomly chosen for deletion, including the internal and terminal regions of the chromosome (Table 1). The genome regions were deleted individually or combined. Twelve strains were generated, including two previously reported strains. The integrant strains were identified by diagnostic PCR, in which the deletion cassettes were assembled upstream or downstream of the target sequence. Target sequence and selective marker were excised without scar after the induction of mazF, and the excision was verified by PCR (Fig. S1, Supplementary data).
Growth profiles of deletion strains revealed genes related to oxygen response
To evaluate the effects of deleting specific chromosomal regions on growth, we investigated the growth profile of mutant strains in liquid YPD medium. Cells of the twelve viable deletion strains and the parent strain were cultivated in YPD medium at 30 °C under aerobic and anaerobic conditions. NK11, NK12, and NK13 showed equal or better growth rates compared with BY4741 under both conditions (Figs. 1A and 1B). By contrast, NK34, NK42 and NK43 grew slightly slower than the wild type; this finding was expected, because the deleted regions consisted of non-essential genes. Interestingly, under aerobic conditions, six strains with deleted YOR124C/YOR134W or YIL014W/YIL005W region (strains NK32, NK33, NK38, NK39, NK44, and NK45) showed higher growth deficiency than other strains when compared with the wild type BY4741 (Fig. 1A). These six strains showed similar growth rates both under aerobic and anaerobic conditions, whereas the other strains and the wild type showed higher growth rates under aerobic than anaerobic conditions (Fig. 1B, 1C). Thus, some of the gene(s) missing in these two genome fragments are needed for aerobic metabolism to function completely.
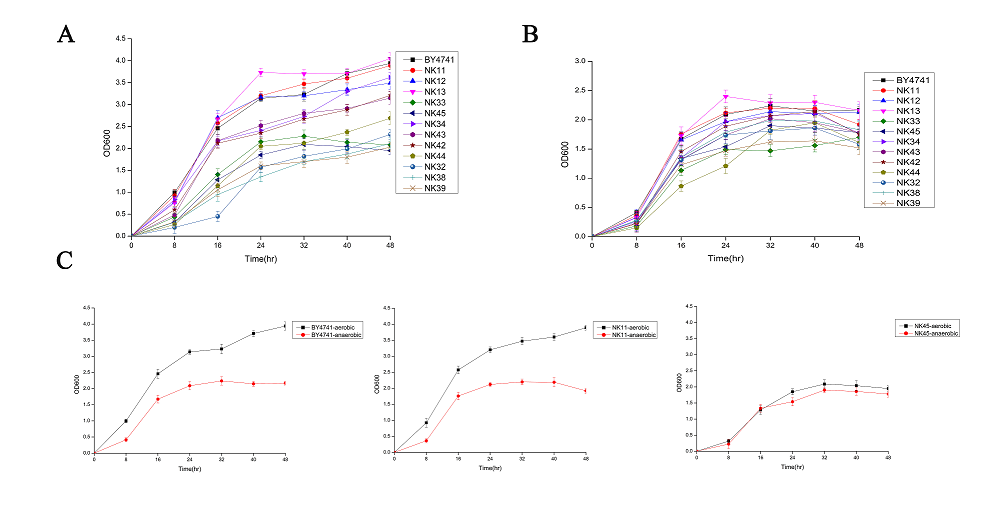
Fig. 1. Growth graphs of deletion strains. (A) Growth profiles of large genome deletion strains and wild type strain BY4741 under aerobic conditions. (B) Growth profiles of large genome deletion strains and wild type strain BY4741 under anaerobic conditions. (C) Growth profiles contrasting the growth under aerobic and anaerobic conditions for wild type strain BY4741 and mutant strains NK11 and NK45. NK11 represents the fast growers in aerobic conditions and NK45 represents the slow growers in aerobic conditions. The mutant strain names are shown in Table 1.
Stress resistance test revealed genes related to ethanol tolerance and high salinity sensitivity
To determine whether the deletion strains exhibited changes in stress sensitivity, we analyzed phenotypic behavior of the deletion strains under several physiological conditions, including heat shock, high salinity, 8% ethanol,hyperosmotic stress, and alkaline (Fig. 2). Except with 1 M NaCl and 8% ethanol, the growth of all the strains was nearly the same as that observed on the YPD plate, thereby indicating that the deletion of the clusters of genes in all the reduced genome regions was not lethal for these stress conditions. Strains NK34, NK42, and NK43 were sensitive to high salinity stress, thereby indicating that the deleted region YOL020W/YOL011W is necessary for cell growth under high salinity conditions. The deletion of YOL019W caused sensitivity to high salinity conditions in yeast [14]. We repeated the NaCl resistance test using the strains with the respective deletion of the genes included in the genome region (Fig. 3A). The single gene knock-out strains did not show high salinity sensitivity, suggesting that deletion of a group of genes or the gene interactions could be responsible for high salinity sensitivity. It is reported that the synergistic effect caused by the loss of multiple genes may reduce the stress response in E. coli [4], which is consistent with our findings with the deletion yeast strains.

Fig. 2. Stress sensitivity test of deletion mutants. Serial tenfold dilution of the described pre-cultures was spotted onto a solid YPD media and plates were incubated at 30°C for 3 days. Concentrations of used inhibitors were as follows: ethanol, 8%; NaCl, 1M and sorbitol, 1.5 M. For heat-shock response, the cell suspension was incubated at 50 °C for 30 min and cooled to 30°C, followed by spotting on YPD plate. Alkaline (pH 8.5) media were prepared by adding 0.1 M Tris·HCl buffer, pH 8.5, to YPD medium. The experiment was repeated three times and showed a similar tendency.
In particular, the strains NK33 and NK45 (and some other strains, to a lesser extent) showed growth deficiency in the presence of high concentration of ethanol (Fig. 2). In the same way, we tested the ethanol tolerance of single gene deletion strains included in the region YOR124C/YOR134W (Fig. 3B). This phenomenon was due to the absence of the following genes in the region: ADE2 (required for purine nucleotide biosynthesis); CAT5 (required for ubiquinone biosynthesis); and RGA1 (GTPase-activating protein for polarity-establishment protein Cdc42p). Growth analysis under ethanol stress of single gene deletion strains showed that absence of ADE2 and RGA1 could result in ethanol sensitivity [14]. To study this possibility, reconstructed strains were then subjected to ethanol stress test and showed good ethanol tolerance again, whereas the growth of strains with control plasmid was slower (Fig. 4). Each one of the ADE2, CAT5, and RGA1 genes was needed for higher ethanol tolerance.
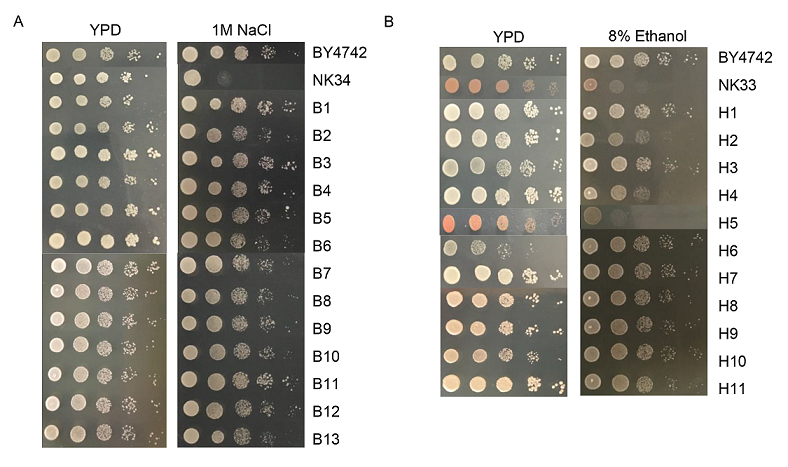
Fig. 3. Stress tolerance tests of single gene deletion strains. (A) High salinity test of strains with the single genes deleted involved in YOL020W/YOL011W. (B) Ethanol tolerance test of strains with the single genes deleted involved in YOR124C/YOR134W. H2, H4 and H5 are strains with CAT5, RGA1 and ADE2 deleted, respectively. More information about the strains were shown in Table 2.

Fig. 4. Ethanol tolerance test of parent strain with control plasmid pLC42, and cat5Δ, rga1Δ and ade2Δ strains complemented with the deleted genes. Cultures were spotted onto SC media with ethanol, except that ade2Δ related strains were spotted onto YPD plate since the synthetic medium contains rich adenine. The experiment was repeated three times and showed a similar tendency.
In this paper, we deleted large genome regions and obtained 12 strains with deletions from 21 kb to 80 kb through the mazF-mediated deletion system. The growth profile and stress sensitivity of these strains were then analyzed. Growth rate analysis showed that the deletion of region YOR124C/YOR134W or YIL014W/YIL005W affected aerobic growth. The yeast showed growth advantage under conditions with oxygen, whereas the mutant strains with one of these two regions showed a similar growth tendency regardless of the presence or absence of oxygen. We assumed that these regions might result in the disability of the oxygen utilization and sensing. The production of heterologous α-amylase in yeast could be improved in anaerobic batch fermentations [15]. In Pichia pastoris, the low oxygen condition could enhance the production of human Fab fragment [16]. Instead of cultivating cells anaerobically, Liu et al. improved heterologous α-amylase secretion at aerobic conditions by activating hypoxia-induced genes in yeast [17, 18] The induction of hypoxia-induced genes or the deletion of HAP1 could induce the switch from aerobic to anaerobic metabolism even in the presence of oxygen [19]. This switch could be an advantage for the production of proteins, such as recombinant human hemoglobin, because it allows the cells to express genes that are normally only expressed under anaerobic conditions and to receive abundant oxygen, which is suitable for heme overproduction. Although the specific genes responsible for this property in mutants with region YOR124C/YOR134W or YIL014W/YIL005W deleted remain unknown, these could be the starting strains for the improvement of the production of heterologous proteins.
Genes whose deletions resulted in ethanol sensitivity were clarified into several functional categories including “aerobic respiration,” “vacuolar transport,” “metabolism of tryptophan,” and “mitochondrion” [14]. Categories of “aerobic respiration” and “mitochondrion” include genes involved in mitochondrial functions, such as ubiquinone (coenzyme Q) biosynthesis (COQ5, COQ9, and COQ10). CAT5 (COQ7) also works for the biosynthesis of coenzyme Q. When cells are exposed to ethanol, S. cerevisiae struggles to use its mitochondrial ETC machinery; therefore, coenzyme Q production becomes essential [20]. This explains the ethanol sensitivity trait of the cat5Δ strain. The deletion of RGA1 affected the function of Cdc42p, which is important for vacuole membrane fusion and could be classified into the functional category of “vesicular transport.” ADE2 serves a function in purine nucleotide biosynthesis. We infer that ethanol enhances the degradation of purine, thereby resulting in the ethanol sensitivity of ade2Δ mutant. Genes could therefore be targets for further genetic modification, which could improve the ethanol tolerance of yeast, especially in the industrial production of ethanol or other metabolites.
Essential genes are defined as genes of an organism that are indispensable for its survival. Although the deletion regions chosen in this study were defined to contain non-essential genes, in the functional assays, we observed that the deleted regions may still have an effect in the cell performance in specific conditions. Organisms have quite much reserve potential in their genetic system to adapt to various conditions. The genes defined as non-essential genes might still have unknown functions, especially in specific conditions. This may also limit the versatility of using minimized artificial chromosomes in synthetic biology.
The authors are grateful to Dr. Li Bingzhi and Pro. Yuan Yingjin from Tianjin University, China, for providing the single gene knock-out strains.
Author contributions
Yuzhen Wu and Quanli Liu designed and performed the experiments, analyzed the data, and completed the manuscript. Yiming Chen, Haijin Xu, Ossi Pastinen, Ossi Turunen, and Mingqiang Qiao discussed the results, commented on the manuscript, and approved the manuscript submission.
Funding
This work was supported by the Tianjin Key Research Program of Application Foundation and Advanced Technology, China (17JCZDJC32200), the Sino-Swiss scientific and technological cooperation project supported by the Ministry of Science and Technology of China (2015DFG32140), and the National Natural Science Foundation of China (31770102).
Supplementary DATA FILE
https://www.siftdesk.org/articles/images/549/s1.pdf
Kellis M, Birren BW, Lander ES. Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae. Nature 2004; 428:617. PMid:15004568
View Article PubMed/NCBITurunen O, Seelke R, Macosko J. In silico evidence for functional specialization after genome duplication in yeast. FEMS Yeast Res 2009; 9:16-31. PMid:19133069
View Article PubMed/NCBIMarsit S, Leducq JB, Durand É, et al. Evolutionary biology through the lens of budding yeast comparative genomics. Nature Reviews Genetics 2017. PMid:28714481
View Article PubMed/NCBIHirokawa Y, Kawano H, Tanaka-Masuda K, et al. Genetic manipulations restored the growth fitness of reduced-genome Escherichia coli. J Biosci Bioeng 2013; 116:52-58. PMid:23477741
View Article PubMed/NCBIMizoguchi H, Sawano Y, Kato J, et al. Superpositioning of deletions promotes growth of Escherichia coli with a reduced genome. DNA Res 2008; 15:277-284. PMid:18753290
View Article PubMed/NCBILi Y, Zhu X, Zhang X, et al. Characterization of genome-reduced Bacillus subtilis strains and their application for the production of guanosine and thymidine. Microb Cell Fact 2016; 15:94. PMid:27260256
View Article PubMed/NCBIMurakami K, Tao E, Ito Y, et al. Large scale deletions in the Saccharomyces cerevisiae genome create strains with altered regulation of carbon metabolism. Appl Microbiol Biotechnol 2007; 75:589-597. PMid:17345083
View Article PubMed/NCBIOhta E, Nakayama Y, Mukai Y, et al. Metabolomic approach for improving ethanol stress tolerance in Saccharomyces cerevisiae. J Biosci Bioeng 2016; 121:399-405. PMid:26344121
View Article PubMed/NCBIPark WK, Yang JW, Kim HS. Identification of novel genes responsible for salt tolerance by transposon mutagenesis in Saccharomyces cerevisiae. J Ind Microbiol Biotechnol 2015; 42:567-575. PMid:25613285
View Article PubMed/NCBILiu Q, Wu Y, Yang P, et al. mazF-mediated deletion system for large-scale genome engineering in Saccharomyces cerevisiae. Res Microbiol 2014; 165:836-840. PMid:25463384
View Article PubMed/NCBIHirashima K, Iwaki T, Takegawa K, et al. A simple and effective chromosome modification method for large-scale deletion of genome sequences and identification of essential genes in fission yeast. Nucleic Acids Res 2006; 34:e11. PMid:16434698
View Article PubMed/NCBIGiaever G, Chu AM, Ni L, et al. Functional profiling of the Saccharomyces cerevisiae genome. Nature 2002; 418:387. PMid:12140549
View Article PubMed/NCBIKnop M, Siegers K, Pereira G, et al. Epitope tagging of yeast genes using a PCR-based strategy: more tags and improved practical routines. Yeast 1999; 15:963. 1097-0061(199907)15:10B<963::AID-YEA399>3.0.CO;2-W
View ArticleYoshikawa K, Tanaka T, Furusawa C, et al. Comprehensive phenotypic analysis for identification of genes affecting growth under ethanol stress in Saccharomyces cerevisiae. FEMS Yeast Res 2009; 9:32-44. PMid:19054128
View Article PubMed/NCBILiu, Zihe, Osterlund, et al. Anaerobic alpha-Amylase Production and Secretion with Fumarate as the;Final Electron Acceptor in Saccharomyces cerevisiae. Applied & Environmental Microbiology 2013; 79:2962-2967. PMid:23435897
View Article PubMed/NCBIBaumann K, Adelantado N, Lang C, et al. Protein trafficking, ergosterol biosynthesis and membrane physics impact recombinant protein secretion in Pichia pastoris. Microbial Cell Factories,10,1(2011-11-03) 2011; 10:93. PMid:22050768
View Article PubMed/NCBIMartínez JL, Liu L, Petranovic D, et al. Engineering the oxygen sensing regulation results in an enhanced recombinant human hemoglobin production by Saccharomyces cerevisiae. Biotechnology and Bioengineering 2015; 112:181-188. PMid:25082441
View Article PubMed/NCBILiu L, Zhang Y, Liu Z, et al. Improving heterologous protein secretion at aerobic conditions by activating hypoxia-induced genes in Saccharomyces cerevisiae. FEMS Yeast Res 2015; 15. PMid:26220688
View Article PubMed/NCBIMartínez JL, Liu L, Petranovic D, et al. Engineering the oxygen sensing regulation results in an enhanced recombinant human hemoglobin production by Saccharomyces cerevisiae. Biotechnology & Bioengineering 2015; 112:181. PMid:25082441
View Article PubMed/NCBIKar A, Beam H, Borror MB, et al. CLD1 Reverses the Ubiquinone Insufficiency of Mutant cat5/coq7 in a Saccharomyces cerevisiae Model System. PLoS One 2016; 11:e0162165. PMid:27603010
View Article PubMed/NCBI