Ana Lúcia Fachin
FAX + 55 -16-36037030
Email: afachin@unaerp.br
© 2019 Sift Desk Journals. All Rights Reserved
VOLUME: 5 ISSUE: 1
Page No: 1-7
Ana Lúcia Fachin
FAX + 55 -16-36037030
Email: afachin@unaerp.br
Gabriela Gonzalez Segura 1, Cyntia Aparecida Montagneri Arevabini 1, Mariana Heinzen de Abreu 1, Tamires Aparecida Bitencourt 1,2, Felipe Garcia Nishimura1, Rene Beleboni 1, Mozart Marins 1, Ana Lúcia Fachin1*
1 Biotechnology Unit, University of Ribeirão Preto, UNAERP, Brazil.
2 Department of Genetics, Ribeirão Preto Medical School, University of São Paulo, Ribeirão Preto, Brazil.
Shufen Wu(wushufen@tust.edu.cn)
Sylwia Zieli%c5%84ska(sylwia.zielinska@umed.wroc.pl)
Taha Majid Mahmood Sheikh(tahamajid1705@yahoo.com)
Jia Yu(yujia_yc@163.com)
Gabriela Gonzalez Segura. The molecular and analytical methods reveal aflatoxin B1-producing Aspergillus flavusisolated from ready-to-eat peanut samples are resistant to the antifungal agent methyl-thiophanate(2019)Journal of Food Science & Technology 5(1) p:1-7
The peanut is a product of high nutritive value and an economically important crop. However, peanuts can be contaminated during storage with Aspergillus flavus, which produces aflatoxins. In humans, aflatoxins can cause hepatocellular carcinoma. Exposure of animals to aflatoxins can result in weight loss and generate residues in meat and milk. The objective of this study was to isolate and identify aflatoxigenic fungi, to quantify the production of aflatoxin B1, and to correlate its production with resistance to the antifungal agent methyl-thiophanate (MT) in ready-to-eat peanuts samples. Nine of the 50 samples selected were found to be contaminated with A. flavus by PCR amplification of the ITS region. In addition, three genes of the aflatoxin B1 biosynthesis pathway were amplified. HPLC analysis revealed that the isolates produced high levels of aflatoxins ranging from 4 to 285 μg/mL. Furthermore, the isolate producing the highest concentration of aflatoxin (285 μg/mL) exhibited the highest resistance to antifungal MT (MIC ≥ 250 µg/mL), factors that might be related. The present results confirm the presence of aflatoxin-producing fungi resistant to MT in peanut samples, which may pose health risks to consumers.
Keywords: HPLC, VER, APA, OMT, mycotoxins, methyl-thiophanate, resistance, antifungal
The peanut (Arachis hypogaea L.) is a legume native to South America and the most widely grown crop around the world because of its pleasant taste and high content of proteins with high biological value, a fact that renders it a product for raw consumption (TOOMER, 2017).
In Brazil, the export of peanuts has increased from 9% in 2016 to 61% in 2017, demonstrating that the development of this crop is extremely important for the Brazilian economy, as well as for the world economy Sampaio; Mogiana (2018). However, peanuts are susceptible to contamination with fungi such as Aspergillus flavus, A. parasiticus and A. nomius. In addition to peanut, these fungi infect other foods such as corn, peppercorn, pistachio, chestnut, walnut and products derived from these grains, which are high-value crops in the international market (KRTZMAN et al., 1987; IDRIS et al., 2010; ALDARS-GARCIA et al., 2018).
Aspergillus flavus produces a carcinogenic metabolite called aflatoxin (OHKURA et al., 2018). About 20 different types of aflatoxins have been identified and the most common are aflatoxins B1, B2, G1 and G2. Studies have shown that A. flavus produces aflatoxins B1 and B2, with the former being the most toxic (LEE et al.1986; JAIMEZ et al. 2000). Peanuts can be contaminated with these mycotoxins during harvest, storage, processing, and distribution (KADER; HUSSEIN, 2009).
Aflatoxin is converted to aflatoxin-8,9-epoxid in the liver of humans or animals. In humans, the effects caused by ingestion of this mycotoxin can be chronic or acute, including hepatotoxic and immunotoxic effects, an increase in reactive oxygen species, and modifications in DNA strands and protein structure (EATON; GALLAGHER, 1994).
Crops such as corn, soybean, peanut and wheat are commonly used in animal feed and have become vehicles of aflatoxin-producing fungi (RUSTOM, 1997). In general, animals that feed on contaminated grains store the mycotoxin in the liver, with consequent health risks such as weight loss, liver damage and kidney problems (QUEZADA et al., 2000). A strong relationship exists between the presence of aflatoxin in dairy cattle ration and the presence of this mycotoxin in milk used for human consumption (OMEIZA et al., 2018).
The objective of this study was to isolate and identify aflatoxigenic fungi in ready-to-eat peanut samples collected in the State of São Paulo, Brazil, as well as to quantify the production of aflatoxin B1 and to correlate this production with resistance to the antifungal agent methyl-thiophanate (MT).
2.1 Isolation, identification and antifungal activity of aflatoxin-producing fungi
Aspergillus flavus ATCC 9643, obtained from Fundação André Tosello and used as control, was cultured on Sabouraud-dextrose agar (Oxoid, Hampshire, England) for 4 days at 35°C according to CHAI et al. (2009). The isolation of the aflatoxin-producing fungi and the determination of antifungal activity were previously described by AREVABINI et al. (2014). Aspergillus flavus was isolated from peanut samples collected at different retail markets in the region of Ribeirão Preto, São Paulo, Brazil, according to SHAPIRA et al. (1996), with described below: the peanut kernels were washed in sterile water and incubated on Czapeck medium (Merck, Germany) for 7 day at 35°C. The A. flavus isolates were then streaked on Czapeck medium to obtain pure cultures.
2.2 DNA extraction
Once pure cultures were obtained, DNA was extracted with TRIzol Reagent® following manufacturer’s recommendations and agarose gels were prepared according to of SHAPIRA et al. (1996), with modifications described below:mycelia were collected by scraping the surface of the colonies with a sterile spatula and inoculated into 100 mL liquid Sabouraud medium (Oxoid, Hampshire, England). The cultures were incubated for 24 h at 35°C under shaking at 150 rpm. After incubation, the mycelia were filtered through a Buchner funnel and pressed between sheets of filter paper to remove excess water. The mycelia were immersed in liquid nitrogen and ground in a sterile mortar and pestle. About 100-150 mg of mycelium was homogenized in 1 mL extraction buffer (200 mM Tris-HCl, pH 8.5, 250 mM NaCl, 25 mM EDTA, 0.5% SDS).
2.3 Detection of aflatoxigenic Aspergillus flavus by the PCR technique
After the extraction of genomic DNA from the isolates, PCR was performed using the primers ITS18S and ITS28S, as well as primers targeting the omt-1, ver-1 and apa-2 genes (SHAPIRA et al., 1996; ZACHOVA et al., 2003; LEVIN, 2012). The PCR assays were conducted as described by SHAPIRA et al. (1996). The primers used are shown in Table 1.
Table 1. Sequence of the primers used for the detection of aflatoxin-producing fungi
|
Primer |
Sequence |
Reference |
|
VER-496 VER-1391 |
5’-ATGTCGGATAAATCACCGTTTAGATGGC-3’ (F) 5’-CGAAAAGCGCCACCATCCACCCCAATG-3’ (R) |
Shapira et al., 1996 |
|
APA-450 APA-1482 |
5’-TATCTCCCCCCGGGCATCTCCCGG-3’ (F) 5’-CCGTCAGACAGCCACTGGACACGG-3’(R) |
Shapira et al., 1996 |
|
OMT-208 OMT-1232 |
5’-GGCCCGGTTCCTTGGCTCCTAAGC-3’ (F) 5’-CGCCCCAGTGAGACCCTTCCTCG-3’ (R) |
Shapira et al., 1996 |
|
ITS 1 (18S) ITS 4 (28S) |
5’-TCCGTAGGTGAACCTGCGG-3’ (F) 5-TCCTCCGCTTATTGATATG-3’(R) |
White et al., 1990 |
2.4 Purification and sequencing of the PCR products
The PCR-positive samples were purified using the GFX Purification Kit (GE Healthcare, USA) following manufacturer’s recommendations. The PCR products were directly sequenced using the ITS 1 and ITS 4 primers and DNA sequencing was performed on a Perkin-Elmer/ABI model 373 DNA Sequencer.
2.5 Fermentation conditions for aflatoxin B1 production
In 250-ml Erlenmeyer flasks, 1 x 106 conidia/mL were added to 50 mL YES fermentation medium and incubated for 10 days under moderate shaking at 27ºC in the dark, according to CVETNIC and PEPELJNJAK (2007).
2.6 High-performance liquid chromatography (HPLC/UV-DAD)
For the quantification of aflatoxin production by each isolate, a standard curve was constructed by HPLC using aflatoxin B1 (Sigma). HPLC analysis was carried out on a Shimadzu chromatograph (LC-10AD vp) coupled to a diode array detector using a Phenomenex C18 column (25 cm x 4.6 mm, id 5 μm). The mobile phase consisted of water:methanol:acetonitrile (50:40:10), eluted a flow rate of 0.80 mL.min-1. Twenty microliter of each sample was injected and the analyses were monitored at a wavelength of 350 nm.
3.1 Isolation of aflatoxin-producing Aspergillus flavus strains
Fifty peanut samples collected at different retail markets in the region of Ribeirão Preto from June 2011 to October 2012 were analyzed. Nine of the 50 samples exhibited the growth of Aspergillus flavus strains.
3.2 Identification of Aspergillus flavus isolates and of the genes involved in aflatoxin biosynthesis by PCR
PCR permitted the detection of a band corresponding to a 694-bp fragment of the ITS region in the selected isolates. This band is characteristic of A. flavus (HENRY et al., 2000). In addition, the amplification pattern observed for the ATCC strain was similar to that of the isolates (Figure 1).
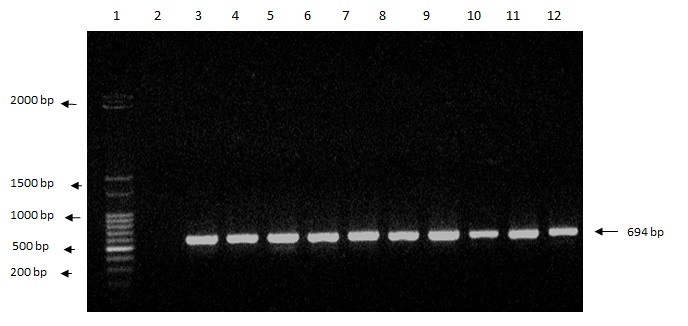
Figure 1. Agarose gel (1.2%) electrophoresis showing amplification of the 694-bp fragment of the ITS gene that characterizes Aspergillus flavus. Lanes: 1: 1-kb marker; 2: blank; 3: isolate Afl1; 4: isolate Afl2; 5: isolate Afl3; 6: isolate Afl5; 7: isolate Afl6; 8: isolate Afl7; 9: isolate Afl8; 10: isolate Afl9; 11: isolate Afl10; 12: A. flavus ATCC 9643.
We also observed amplification (expression) of the apa-2 (a regulator of aflatoxin biosynthesis, which controls the expression of the ver-1 and nor-1 genes), omt-1 (the gene encoding sterigmatocystin-o-methyltransferase), and ver-1 genes (versicolorin A dehydrogenase) (Figures 2, 3 and 4) (SHAPIRA et al., 1996).
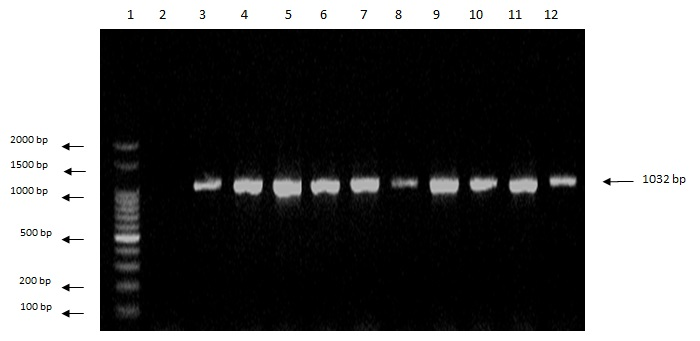
Figure 2. Agarose gel electrophoresis showing amplification of a 1032-bp fragment of the apa-2 gene. Lanes: 1-kb marker; 2: blank; 3: isolate Afl1; 4: isolate Afl2; 5: isolate Afl3; 6: isolate Afl5; 7: isolate Afl6; 8: isolate Afl7; 9: isolate Afl8; 10: isolate Afl9; 11: isolate Afl10; 12: A. flavus ATCC 9643.
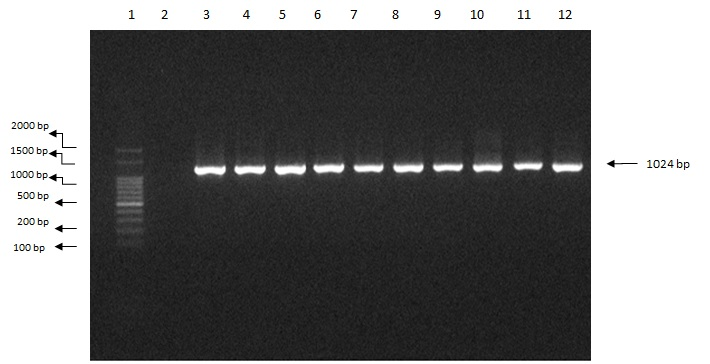
Figure 3. Agarose gel electrophoresis showing amplification of a 1024-bp fragment of the omt-1 gene. Lanes: 1: 1-kb marker; 2: blank; 3: isolate Afl1; 4: isolate Afl2; 5: isolate Afl3; 6: isolate Afl5; 7: isolate Afl6; 8: isolate Afl7; 9: isolate Afl8; 10: isolate Afl9; 11: isolate Afl10; 12: A. flavus ATCC 9643.
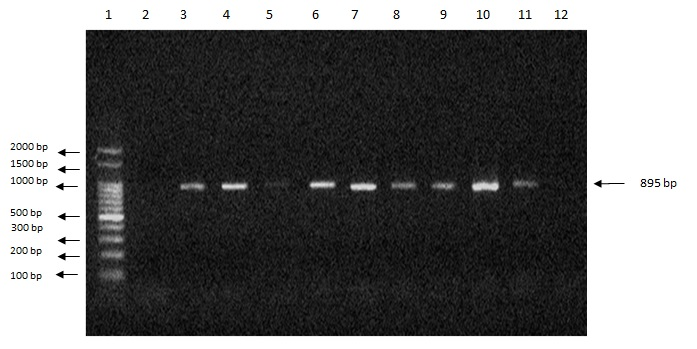
Figure 4. Agarose gel electrophoresis showing amplification of an 895-bp fragment of the ver-1 gene. Lanes: 1: 1-kb marker; 2: blank; 3: isolate Afl1; 4: isolate Afl2; 5: isolate Afl3; 6: isolate Afl5; 7: isolate Afl6; 8: isolate Afl7; 9: isolate Afl8; 10: isolate Afl9; 11: isolate Afl10; 12: A. flavus ATCC 9643.
All A. flavus isolates obtained from peanuts as well as the A. flavus ATCC 9643, showed similarity to strains deposited in the database of the National Center for Biotechnology Information (NCBI). Amplification of the ITS gene revealed identity with ITS regions of the ribosomal 18S and 28S gene of A. flavus (accession number JF951750.1). Likewise, analysis of samples for the omt-1 gene revealed identity with the O-methyltransferase gene of an A. flavus isolate (accession number L25836.1) and the ver-1 gene showed identity with the reported gene of an A. flavus strain (accession number AB007805).
3.3 Quantification of aflatoxin B1 by HPLC/UV-DAD and antifungal activity
The detection and quantification of aflatoxin B1 by HPLC/UV-DAD demonstrated that 7 fungi samples were aflatoxin producers. Afl7 was the isolate with the most expressive production of the mycotoxin, followed by Afl10, Afl9 and Afl2. No aflatoxin production was also observed for the ATCC strain. In this study, 4 of the A. flavus isolates were resistant to MT compared to the MIC of 3.9 µg/mL of the A. flavus ATCC strain (Table 2).
Table 2. Concentration of aflatoxin B1and determination of Minimal Inhibitory Concentration (MIC) in (µg/mL) of Aspergillus flavus strains
|
Strain
|
Concentration of aflatoxin B1 |
MIC of methyl-thiophanate (MT)
|
||||
|
ATCC |
ND |
3.9 |
|
|||
|
Afl1 |
18 ± 1 |
7.8 |
|
|||
|
Afl 2 |
135 ± 5 |
7.8 |
|
|||
|
Afl3 |
47 ± 1 |
3.9 |
|
|||
|
Afl5 |
4 ± 1 |
3.9 |
|
|||
|
Afl6 |
ND |
15 |
|
|||
|
Afl7 |
285 ± 11 |
≥ 250 |
|
|||
|
Afl8 |
ND |
1.9 |
|
|||
|
Afl9 |
168 ± 6 |
1.9 |
|
|||
|
Afl10 |
193 ± 7 |
1.9 |
|
|||
ND: not detectable.
Despite hygiene-sanitary control of grains destined for human consumption, nine of the 50 peanut samples collected in the present study were contaminated with A. flavus, the main producer of aflatoxin B1.
The present study confirmed the presence of A. flavus in 18% of the peanut samples by the amplification of bands corresponding to the ITS18S and ITS28S genes. Mahmoud (2015) identified A. flavus in 53.6% (n=28) of peanut samples collected in Saudi Arabia by PCR amplification of the FL1 gene.
Comparison of the molecular and analytical (HPLC/UV-DAD) methods showed that both were adequate for detecting aflatoxin in A. flavus isolates. Aflatoxin B1 was quantified by HPLC in fungal isolates with positive PCR results for aflatoxin biosynthesis genes (omt-1, ver-1 and apa-2). Aflatoxin B1 levels ranging from 4 to 285 µg/mL. Mahmoud (2015) detected aflatoxin B levels ranging from 1.64 to 109.18 µg/mL by HPLC in 70.37% of A. flavus strains isolated from peanuts. Mupunga et al. (2014) also observed contamination with A. flavus and A. parasiticus in peanut and peanut butter samples collected in Zimbabwe. Oliveira et al. (2009) selected 240 peanut samples in Brazil and found positive results for aflatoxins (B1, B2, G1, and G2) in 44.2%.
The isolate Afl7 produced the highest level of aflatoxin B1 (285 µg/mL) and was resistant to MT (MIC ≥ 250 µg/mL) as shown by Arevabini et al. (2014). Moreover, the isolate Afl2 produced high levels of aflatoxin B1 (135 µg/mL) and was also resistant to MT (MIC of 7.8 µg/mL). These fungal isolates may have been selected by the indiscriminate use of this commercial antifungal agent, which persisted in the field because of their resistance.
Methyl-thiophanate is a fungicide widely used in global agriculture for the control of many fungal diseases in vegetable, fruit and field crop plantations IBTISSEM et al., (2017).
The problem of the abusive use of pesticides in agriculture is linked to the agricultural production policies in Brazil, which are based on quantity rather than quality of the food and whose focus is unprecedented productivity gain (PIGNATI et al., 2017).
To guarantee the food and nutritional safety of the population, control parameters needs to be established for the effective monitoring of pesticide use, as well as incentives for the development of natural antifungal agents that pose no public health risks. AREVABINI et al. (2014) demonstrated that marine natural products were more effective against MT-resistant A. flavus isolates.
Resistance to MT could be related to aflatoxin production. These data makes us question the efficiency of the antifungal agents used in peanuts for elimination of aflatoxigenic fungi in the field t
Isolates Afl6 and Alf8 were found to be resistant to MT, with MIC of 15 µg/mL [16]. However, no aflatoxin B1 production was observed in these isolates. This finding might be due to the detection limit of the analytical technique used, which is on average 2 µg/mL (ESPINOSA-CALDERON et al., 2011). In this case, PCR is a more sensitive technique since it can detect very low amounts of DNA such as nanograms and femtograms (AREVABINI et al., 2014). In addition, A. flavus isolate Afl6 and Af18 may produce a mycotoxin other than aflatoxin B1, such as B2, G1 and G2 (GHAFI et al., 2018)
High levels of aflatoxins in grains are harmful to human and animal health, with a negative impact on the world economy. JAGER et al. (2016) quantified high levels of aflatoxin M1 in urine samples of patients, confirming the consumption of contaminated foods. AMIRKHIZI et al. (2015) observed aflatoxins at relatively high levels in the liver and eggs of chickens. KANG'ETHE AND LANG'A (2009) found contamination of dairy cattle feed with aflatoxins and a consequent expressive increase of aflatoxin B1 and M1 levels in the milk consumed by the population.
Despite sanitation and hygiene control in grain marketing, our study demonstrated the contamination of peanut samples with aflatoxin-producing A. flavus by molecular and analytical methods. In addition, we detected high levels of aflatoxin in two A. flavus isolates that were resistant to the antifungal agent MT. This finding demonstrates the inefficacy of currently used microbiological and chemical control methods.
We thank the staff of Unaerp.
Funding: This study was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, 2016/22701-9) and CAPES (PhD fellowship granted to MHA).
Aldars-García, L., Marín, S., Sanchis, V., Magan, N., & Medina, A. (2018). Assessment of intraspecies variability in fungal growth initiation of Aspergillus flavus and aflatoxin B1production under static and changing temperature levels using different initial conidial inoculum levels. International Journal of Food Microbiology 272:1-11. PMid:29482078
View Article PubMed/NCBIAmirkhizi, B., Arefhosseini, S. R., Ansarin, M., & Nemati, M (2015) Aflatoxin B1in eggs and chicken livers by dispersive liquid-liquid microextraction and HPLC. Food Additives and Contaminants: Part B Surveillance 8(4): 245-249 PMid:26160230
View Article PubMed/NCBIArevabini, C., Crivelenti, Y. D., Abreu, M. H. De, Bitencourt, T. A., Berlinck, R. G. S., Hajdu, E (2014). Antifungal Activity of Metabolites from the Marine Sponges 1-4.
Chai, L. Y. A., Kullberg, B. J., Vonk, A. G., Warris, A., Cambi, A., Latgé, J. P., … Netea, M. G,. Modulation of toll-like receptor 2 (TLR2) and TLR4 responses by Aspergillus fumigatus (2009). Infection and Immunity 77(5):2184-2192. PMid:19204090
View Article PubMed/NCBICvetnić, Z., & Pepeljnjak, S,. Interaction between certain moulds and aflatoxin B1producer Aspergillus flavus NRRL 3251 (2007) Arhiv Za Higijenu Rada i Toksikologiju 58(4):429-434 PMid:18063528
View Article PubMed/NCBIEaton, D. L., & Gallagher, E. P. Mechanisms of aflatoxin carcinogenesis (1994). Annual Review of Pharmacology and Toxicology 34(1):135-172. PMid:8042848
View Article PubMed/NCBIEspinosa-Calderon, A., Miguel, L., Francisco, R., Roberto, J., Guevara Gonzalez, R. G., & Torres-Pacheco, I. Methods for detection and quantification of aflatoxins (2011). Aflatoxins - Detection, Measurement and Control.
View ArticleGhafi, G.O; Safarzad, M; Moazeni, M; Niknejad, F. Quantification of aflatoxin B1, B2, G1, G2 and detection of aflR, aflJ genes in Aspergillus flavus contaminated corn in poultry in Northeast of Iran (2018). Sydowia 70:179-184.
Henry, T., Iwen, P. C., Hinrichs, S. H., Henry, T., Iwen, P. C., & Hinrichs, S. H,. Identification of Aspergillus species using internal transcribed spacer regions 1 and 2 (2000). Journal of clinical microbiology 38(4):1510-1515.
Ibtissem, B. A., Hajer, B. S., Ahmed, H., Awatef, E., Choumous, K., Ons, B., Najiba, Z. Oxidative stress and histopathological changes induced by methylthiophanate, a systemic fungicide, in blood, liver and kidney of adult rats (2017) African Health Sciences 17(1):154-163 PMid:29026389
View Article PubMed/NCBIIdris, Y. M. A., Mariod, A. A., Elnour, I. A., & Mohamed, A. A,. Determination of aflatoxin levels in Sudanese edible oils (2010) Food and Chemical Toxicology. 48(8-9):2539-2541 PMid:20478351
View Article PubMed/NCBIJager, A. V., Tonin, F. G., Baptista, G. Z., Souto, P. C. M. C., & Oliveira, C. A. F. Assessment of aflatoxin exposure using serum and urinary biomarkers in São Paulo, Brazil: A pilot study (2016) International Journal of Hygiene and Environmental Health 219(3):294-300 PMid:26740158
View Article PubMed/NCBIJaimez, J., Fente, C. ., Vazquez, B. ., Franco, C. ., Cepeda, A., Mahuzier, G., & Prognon, P,. Application of the assay of aflatoxins by liquid chromatography with fluorescence detection in food analysis (2000) Journal of Chromatography A., 882(1-2):1-10 00212-0
View ArticleKader, A. A., & Hussein, A. M,. Project on the Development of Sustainable Date Palm Production Systems in the GCC countries of the Arabian Peninsula (2009) HARVESTING AND POSTHARVEST HANDLING OF DATES 1-18.
Kang'Ethe, E. K., & Lang'A, K. A. Aflatoxin B1 and M1 contamination of animal feeds and milk from urban centers in Kenya (2009) African Health Sciences 9(4):218-226
Kurtzman, C. P., Horn, B. W., & Hesseltine, C. W. Aspergillus nomius, a new aflatoxin-producing species related to Aspergillus flavus and Aspergillus tamarii (1987) Antonie van Leeuwenhoek 53(3):147-158. PMid:3116923
View Article PubMed/NCBILevin, R. E,. PCR detection of aflatoxin producing fungi and its limitations (2012). International Journal of Food Microbiology 156(1):1-6. PMid:22445201
View Article PubMed/NCBIMahmoud, M. A. Detection of Aspergillus flavus in stored peanuts using real-time PCR and the expression of aflatoxin genes in toxigenic and atoxigenic A. flavus isolates (2015) Foodborne Pathogens and Disease 12(4):289-296. PMid:25621617
View Article PubMed/NCBIMupunga, I., Lebelo, S. L., Mngqawa, P., Rheeder, J. P., & Katerere, D. R. Natural occurrence of aflatoxins in peanuts and peanut butter from Bulawayo, Zimbabwe (2014) Journal of Food Protection 77(10):1814-1818. PMid:25285504
View Article PubMed/NCBIOhkura, M., Cotty, P. J., & Orbach, M. J,. Comparative Genomics of Aspergillus flavus S and L Morphotypes Yield Insights into Niche Adaptation (2018) G3:Genes|Genomes|Genetics 25
View ArticleOliveira, C. A. F., Gonçalves, N. B., Rosim, R. E., & Fernandes, A. M. Determination of aflatoxins in peanut products in the northeast region of São Paulo, Brazil (2009) International Journal of Molecular Sciences 10(1):174-183. PMid:19333440
View Article PubMed/NCBIOmeiza, G. K., Kabir, J., Kwaga, J. K. P., Kwanashie, C. N., Mwanza, M., & Ngoma, L,. A risk assessment study of the occurrence and distribution of aflatoxigenic Aspergillus flavus and aflatoxin B1 in dairy cattle feeds in a central northern state, Nigeria (2018) Toxicology Reports 846-856. PMid:30151345
View Article PubMed/NCBIPignati, W. A., Lima, F. A. N. de S. e, Lara, S. S. de, Correa, M. L. M., Barbosa, J. R., Leão, L. H. da C., & Pignatti, M. G.Distribuição espacial do uso de agrotóxicos no Brasil: uma ferramenta para a Vigilância em Saúde (2017) Ciência & Saúde Coletiva 22(10):3281-3293 PMid:29069184
View Article PubMed/NCBIQuezada, T., Cuéllar, H., Jaramillo-Juárez, F., Valdivia, A. G., & Reyes, J. L,. Effects of aflatoxin B1 on the liver and kidney of broiler chickens during development (2000) Comparative Biochemistry and Physiology - C Pharmacology Toxicology and Endocrinology 125(3):265-272 00107-3
View ArticleRustom, Y. S. Aflatoxin in food and feed: occurrence, legislation and inactivation by physical methods (1997) Food Chemistry 59(1):57-67. 00096-9
View ArticleSampaio, A.; Mogiana, A, 2018. Amendoim : exportações do grão em expansão.
Shapira, R., Paster, N., Eyal, O., Menasherov, M., Mett, A., & Salomon, R, Detection of aflatoxigenic molds in grains by PCR (1996) Applied and Environmental Microbiology 62(9):3270-3273.
The Observatory of Economic Complexity. Peanuts. 2016. Accessed 13 Setember 2018
View ArticleToomer, O. T. Nutritional chemistry of the peanut (Arachis hypogaea) (2017). Critical Reviews in Food Science and Nutrition. Accessed 18 Setember 2018
Zachová, I., Vytřasová, J., Pejchalová, M., Červenka, L., & Tavčar-Kalcher, G,. Detection of aflatoxigenic fungi in feeds using the PCR method (2003) Folia Microbiologica 48(6):817-821 PMid:15058197
View Article PubMed/NCBI