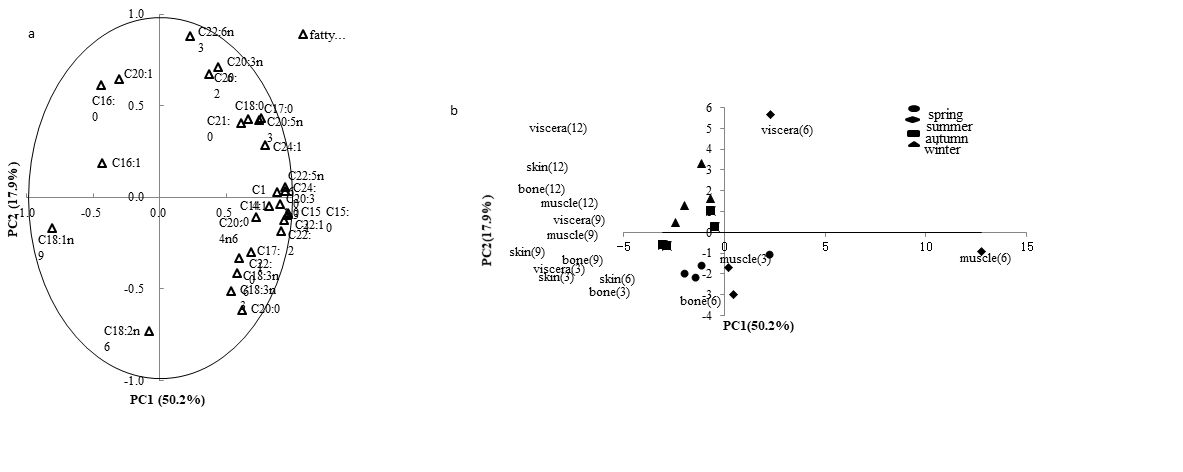
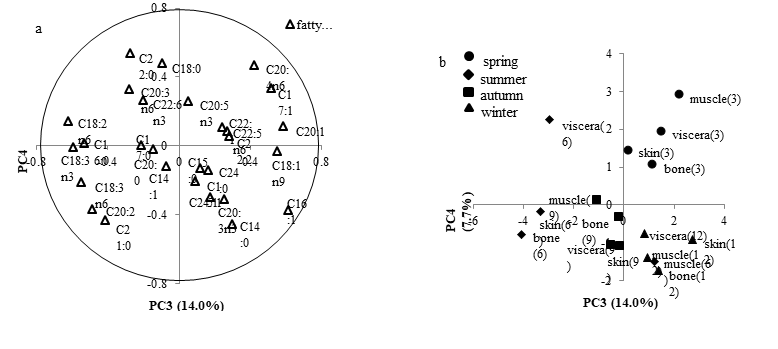
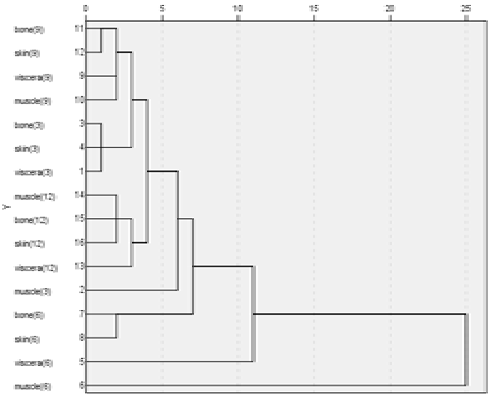
Han Lu, Hui Hong, Yongkang Luo*
College of Food Science and Nutritional Engineering, China Agricultural University, Beijing Higher Institution Engineering Research Center of Animal Product, Beijing 100083 China
*E-mail: luoyongkang@263.net; luoyongkang@cau.edu.cn